Strategic Arts and Sciences Alliance
The Brand Name of the Media Revolution

|
Idiosyntactix Strategic Arts and Sciences Alliance The Brand Name of the Media Revolution |
|
[library]
DCOM - Chapter 5

DEFENDING CYBERSPACE AND OTHER METAPHORS MARTIN LIBICKI
Postcards from the Immune System
Sections
How the Immune System Works The Immune System as an Analog Complex System Some Lessons for Warfare Implications for Information Systems Conclusions
Every living organism is confronted by continual intrusions from its environment. To survive, every organism has therefore had to develop defenses that render it resistant, or immune, to such assaults. These defenses range from physical barriers, s uch as a cell wall, to highly sophisticated systems.[81]
The human immune system has often been explained by metaphors taken from war.[82] In recent years, defense analysts have returned the favor by looking to the immune system for suggestions on how to fight war. The rise of inf ormation warfare helps explain why. First, the defense of large networks against computer viruses and other illicitly entering material may be helped by understanding how the human immune system defends itself against biological viruses.[83] Second, the ability of the immune system to distinguish between self and nonself (i.e., the pathogenic invader) may have parallels in the application of intelligence-based warfare and low-intensity combat (including counter-terrorism and peace operations). Third, complexity theory is gaining attention as a way of explaining warfare, and the immune system is often invoked as a highly functional complex and adaptive system.
If the immune system is to be exploited intelligently as a metaphor it must be understood in and of itself.[84] Its specifics matter, because they are what make the immune system work. To extract certain features from the im mune system in isolation while ignoring the underlying complexities may be to draw analogies to an incomplete system that would not work.[85]
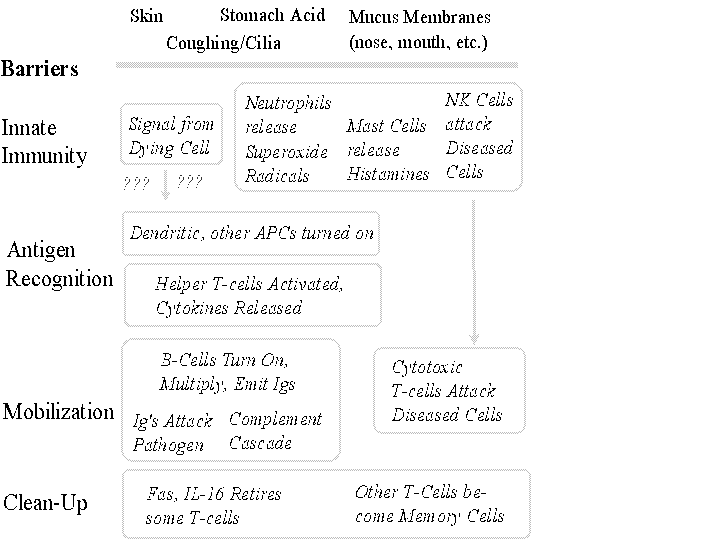
The human body fights a continuous invasion of pathogens from bacteria to viruses, fungi, worms, protozoa, and spirochetes. As Figure 1 illustrates, the human immune defenses entail:
Figure 1
The Immune Response
- barriers,
- innate immune defenses,
- recognition that an antigen is nonself and thus a likely pathogen (or an indicator of pathogenic activity),
- mobilization of biochemical processes to destroy the pathogen (or diseased or cancerous cells),
- returning itself to a standby state.
Like any good defense system, the immune system is multilayered. Skin forms the primary, but not sole, barrier to invasion. Nasal, alimentary and other passages are protected by mucus membranes, which secrete chemicals harmful to bacteria while stomach ac id also is corrosive and cilia and coughing responses protect the lungs.
Because many pathogens can establish an infection despite such barriers, the immune system must be able to detect diseased cells and eliminate them to keep disease from spreading. Yet it must also be careful not to attack healthy cells or destroy its own proteins. This balancing act, which must be maintained over a lifetime, gives the human immune system its daunting complexity.
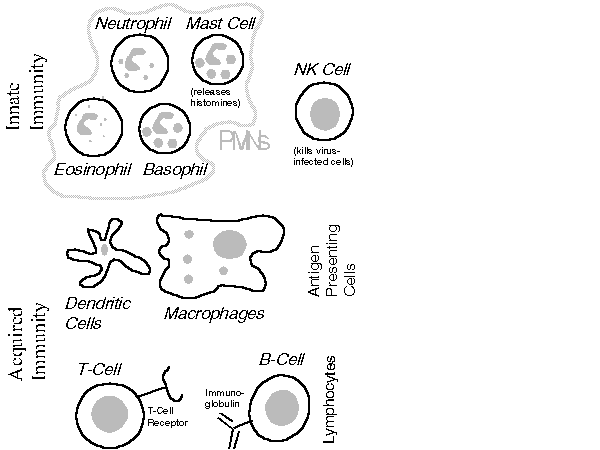
The immune system is generally discussed in terms of innate immunity and acquired immunity. Innate immunity is prompted by specific events (e.g., trauma) and directed against specific invaders that the human immune system has evolved to recognize. Acquired immunity works against nonself antigens which the body has learned to recognize. The key cells of the innate and acquired immune system are portrayed in Figure 2.
The Innate Immune System: Following a cut (or other traumas such as wounds or burns), local platelets in the blood rupture, releasing chemicals that expand the size of blood vessels and raise the local temperature. Both changes induce an influx (ch emotaxis) of polymorphonuclear (PMN) cells whose hydrolytic enzymes, peroxides, and superoxide radicals are toxic to many microorganisms. PMNs (e.g., neutrophils) also clean up dead and dying cells, which otherwise would provide rich feeding grounds for b acteria.
Figure 2
Cells of the Immune System Natural Killer (NK) cells play an early role in combating viral infections that happen to induce the appearance of glycoproteins on the surface of the affected cells. These glycoproteins attract and activate NK cells, which kill infected cells by sending them chemical agents which induce such cells to commit suicide but not release their contents -- apoptosis.
The innate immune system starts work within thirty minutes to an hour after infection and works well against pathogens that elicit specific chemical reactions, but it does not otherwise distinguish between an invader and the body -- that is the job of the acquired immune system, which takes longer (circa two days) to respond but is far more precise.
Recognizing Nonself: The key to the acquired immune system is recognizing pathogens (and related sugar groups) as different from self-proteins so as to attack only the former. Specific lymphocytes (T-cells and B-cells) are what recognize individual pathogens -- more precisely, not the pathogen itself but its antigens[86] (surficial proteins or proteins that result from cellular processing) which, in turn, are recognized by their epitopes (sequences of five to seven am ino acids). Recognition results from a lock-and-key match between a receptor on a T-cell or the immunoglobulin (Ig) molecule of the B- cell.
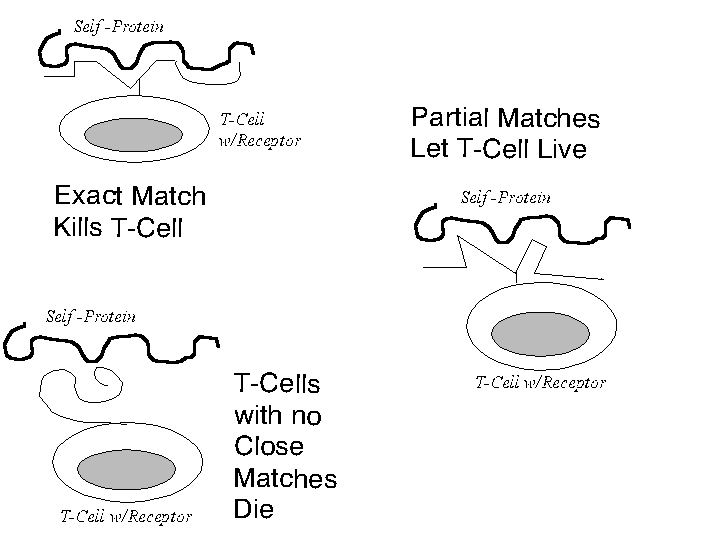
Unfortunately, there are hundreds of thousands of potential antigens in the world. The human genome is too small to code a separate receptor to recognize each antigen. Even if it could, the body would still be defenseless against something humans have nev er encountered. The body, it turns out, does not code a separate receptor for every potential epitope. Instead, during fetal development, a basic toolkit of receptor components is reshuffled to produce up to a hundred million potential receptor combinatio ns; then the body gets rid of those it does not want. How? In the fetal thymus, receptors constantly come in contact with the body's proteins. Lymphocytes whose receptors match the body's own proteins too well have to be discarded because they could induc e an autoimmune reaction later in life[87]. Lymphocytes whose receptors fail to come close to matching the body's own proteins die by neglect probably because they would not recognize any antigen. The survivors (less than on e in ten) are those that exhibit a partial match with the body's own proteins. This threefold differentiation is illustrated in Figure 3.
Figure 3
Clonal Selection of Lymphocytes
The downside of the body's producing millions of different T-cell receptors is that only one in roughly ten thousand T-cells can recognize a given antigen.[88] This is a small force arrayed against an antigen designed to rep licate lustfully once it has settled in. Human survival depends on the body's ability to replicate its immune cells to compete.
Initiating an Acquired Immune Response: Almost all acquired immune responses start with an interaction involving an antigen- presenting cell (APC), an antigen, and a helper T-cell.
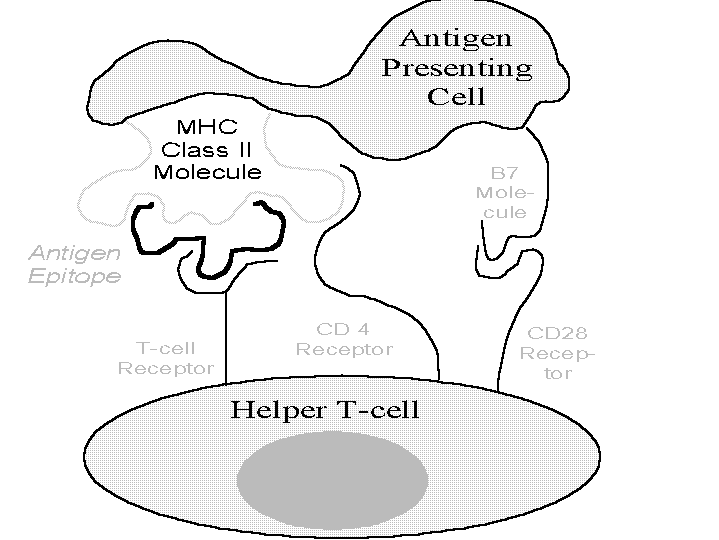
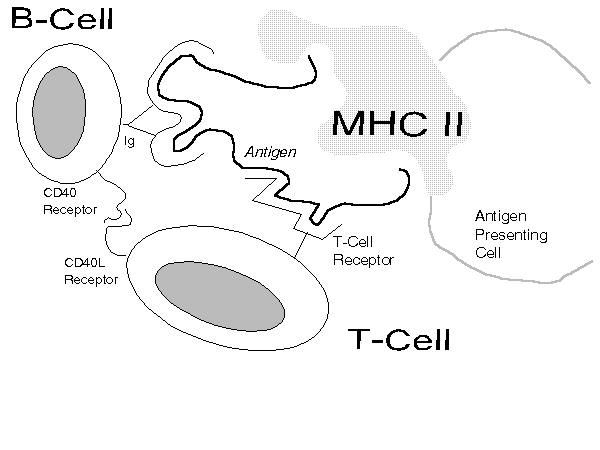
APCs (dendritic cells, macrophages, B-cells) tend to congregate near potential entry points (e.g., skin, lungs, stomach) or in the lymphatic system.[89] These cells can present antigen to T-cells because their surfaces can d isplay major histocompatibility complex[90] (MHC) molecules and co- stimulatory proteins (e.g., the B7 protein). Dendritic cells are particularly rich in both. As Figure 4 illustrates, if there is a close match betwee n the antigen (as held by the MHC molecule) and the T-cell receptor then the T-cell is activated.
Figure 4
T-Cell Recognition of Antigen
MHC molecules are like pliers; they grip segments of antigens (that remain after processing by cellular mechanisms) and present them to T-cells. The shape of an MHC molecule -- and the human body contains a profusion of types -- determines how well it can present specific antigens to T-cells. If T-cells cannot see antigens gripped in a particular way they may not mount a vigorous immune reaction against it. Because the character of the more common pathogenic invaders changes so rapidly, human MHC proteins also have to evolve very rapidly to achieve the best grips. For that reason, populations in certain regions often have unique MHC molecules (e.g., the MHC molecules of those living near the Gambia river are specialized for ambient malaria).
When an antigen so presented achieves a correct lock-and-key match with a helper T-cell's receptor (and certain pairings occur such as those between the T-cell's CD28 receptor and a B7 protein), the helper T-cell is activated.[9 1] Activation (a) causes the helper T-cell to mature and replicate, (b) spit out cytokines (notably interleukins [ILs]) to activate critical effector cells (e.g., cytotoxic T- cells),[92] and (c) act ivate cognate B-cells.
Cytokines exist mostly to stimulate the immune system. Some (IL-1, IL-4, and IL-7) promote T-cell proliferation and excite B-cells. Others (IL-3 and IL-5) affect mast cells and eosinophils. Yet other chemicals attract PMNs and macrophages, which grow incr easingly voracious and will dissolve whatever they ingest (because activation is near an infection site, they are more likely to ingest antigens rather than random cellular material).
Lymphocytes live for a few weeks and then die. A small percentage of these cells does not develop fully and remains in circulation.[93] These undeveloped, longer lived T- cells create a reservoir of potential "memory" cells (roughly five to a hundred times more prevalent than prior to original infection). This population gives the body a head start on a counterattack the next time the antigen appears. Compared to other lymphocytes, memory cells adhere better to dendritic and other APCs and thus react to antigens more quickly. A body invaded once is therefore immune to almost all subsequent invasions[94] of a type memory cells have experienced.
The Role of the B-Cell: The helper T-cell starts the immune response, but does not itself fight the invader. It takes another lymphocyte, such as the B-cell, to release the chemicals that do this. Frequently, an antigen that has attracted a T-cell to one of its many epitopes has also bound B-cells to another of its epitopes. The cognate relationship (see Figure 5) induces replication of B-cells specific to the particular antigen and the release of their attached immunoglobulin[95] (Ig) antibodies.
Figure 5
Cognate Reactions Between B- and T- Cells
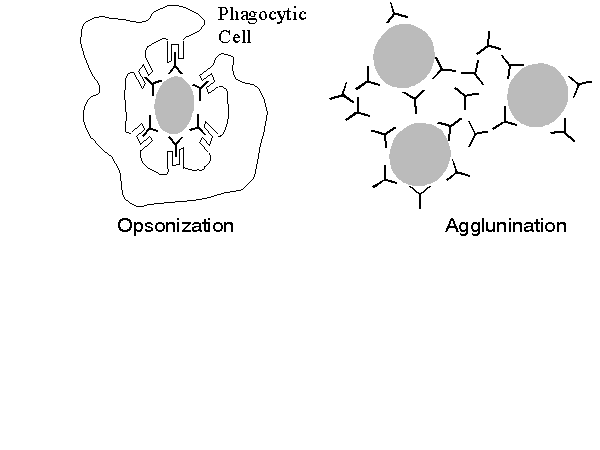
Ig molecules bind to antigens snugly. Opsonization is when a pathogen is coated with enough Ig molecules for a macrophage to grip it like velcro and eat it (smooth-coated bacteria are otherwise hard to grab). Ig-coated pathogens also move slower; when eno ugh Ig (notably IgM) molecules bind enough antigens, the entire mass is agglutinized and immobile. See Figure 6.
Figure 6
How Immunoglobulin Destroys Pathogens
Ig molecules also kill invaders by stimulating[96] the complement reaction, a complex cascade of protein activations and cleavages which yields two final proteins -- one to slice a channel in the antigen and the other to wid en the channel fatally. An intermediate protein in this cascade attracts neutrophils to perform clean-up. The complement reaction can also be turned on by (a) toxins, such as cobra venom, cell walls of gram-negative bacteria, or some yeasts or ( b) by mannose-binding proteins emitted by the liver stimulated by IL- 6. The complement reactions do not distinguish between self and nonself, but since they occur in the immediate vicinity of an antigen-Ig reaction, they are disproportionately aimed at the pathogen, rather than at the body's own tissues.
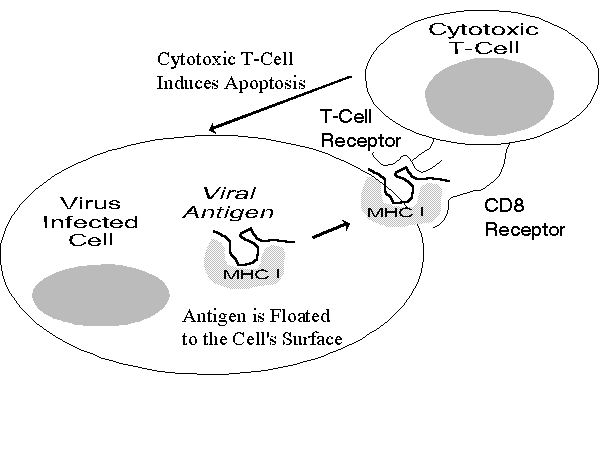
The Role of the Cytotoxic T-Cell: Cytotoxic T-cells are important defenders against viruses. A virus does not stay in circulation very long; after it invades cells it uses the cell's own chemistry for further replication of viral nucleoproteins (e. g., DNA); thus they would normally be inaccessible to the immune system. However, all cells contain MHC class I molecules, which tend to circulate internally and presents protein fragments to the cell's surface. Because cells with viral infections and som e precancerous cells[97] manufacture proteins otherwise not seen in the human body, they can induce immune reactions.
As Figure 7 illustrates, if an antigen brought by a MHC class I molecule encounters a cytotoxic T-cell with the right receptors, the T-cell will inject chemicals into infected cells, which inhibit viral replication and induce apoptosis. Cytotoxic T-cells generally have to be turned on by cytokines; the reaction of a helper T-cell in the neighborhood increases the likelihood that enough cytokines are present to stimulate the cytotoxic T-cell.
Figure 7
Cytotoxic T-Cells
The Immune System as an Analog Complex System
Clausewitz observed that "Everything in war is simple, but the simplest thing is difficult."[98] Similarly, distinguishing between self and nonself is unambiguous but making the distinction in practice is difficult.[99]
The immune system is not perfect. It arose through slow, uncertain, and random processes of evolution.[100] Evolved systems tend to be robust and well tested, but they do not start from scratch. New structures and chemical pathways are usually variations on earlier ones; traits are not discarded but written around or over (so that a feature that no longer makes sense but is seldom harmful is likely to stay in the gene pool). Biochemical mechanisms may be efficient but hardl y precise or deterministic information processors; they depend on the statistical mechanics that result from the random peregrinations of molecules. The human immune system must be sufficiently redundant to withstand most genetic errors and be able hold i ts own against pathogens, whose survival depends on outwitting immune systems.
It important for the human immune system to turn itself off as well as on. Anyone who has suffered from swelling (tumor), redness (rubor), heat (calor), and pain (dolor) intuitively understands why the immune system cannot be kept on all the time. As note d, complement reactions, macrophages, and PMNs tend to attack everything in their immediate vicinity. Every new year brings further evidence that autoimmune disorders are more common than earlier believed, as in lupus, multiple sclerosis, and rheumatoid a rthritis. Diabetes and Alzheimer's disease may also have autoimmune components.
How does the immune system turn itself off? To start with, most of its active components have lifespans measured in weeks; thereafter, memory cells alone persist in semi-active state. The protein fas is used to deactivate and retire mature T-cells. Small resting B-cells may limit T-cell activation by presenting antigens to them but without the B7 molecule.[101] IL-16, secreted by cytotoxic T-cells, may help get rid of infected helper T-cells.[102] Some research suggests the existence of "veto" cells capable of turning off or deleting T cells that recognize antigens on the "veto" cell surface.
Good and bad are closely interwoven in the immune system. A mast cell attached to the stem of an IgE molecule with an antigen in its grip will release tumor necrosis factor alpha, and histamines, both thought to be important in fighting protozoa. Histamin es accelerate the entry of immune cells into the bloodstream and attract PMNs, which also attack invaders. Yet excess histamine production can induce allergic reactions, sometimes leading to anaphylaxis, which can be fatal. Some superantigens are believed to be able to overstimulate T-cells, that is, to produce IL-2 in amounts that can disable or kill.[103] Toxic-shock syndrome and septic-shock syndrome are related to excess cytokine production.
One source of complexity is that pathogens practice, what is in effect, information warfare. Viruses often change protein coats to become unrecognizable to activated immune cells primed for the original signature. The strategy of the AIDS virus is to atta ck the immune system directly, by targeting the helper T-cell as its host. Rapid mutation allows the AIDS virus to present the slowly declining population of helper T-cells with one random antigen after another, until one evokes only a weak immune respons e. Other viruses inhibit MHC I molecules and keep them from appearing, antigen in grip, on the surface of the cell. A pathogen can inhibit an immune system reaction by releasing a chemical that fits a key receptor in the immune process in such a way that the cell fails to turn on; if enough receptors are occupied the immune reaction is minimized -- a form of deception. Other viruses capture and thus turn on genes that produce the proteins that inhibit certain cytokine cascades that would otherwise lead to cell activation.
The problems of immunological defense and national defense differ of course. Nevertheless, it may be instructive to speculate (within limits[104]) on lessons the former might hold for the latter if they were su fficiently similar.
March to the Sound of Guns. Chemotaxis is the method by which components of the innate immune system make their way to the invasion site and thereby respond within minutes and seconds. It holds the fort, so to speak, until the acquired immune syste m can be brought up to speed.
But Shape the Battlefield Soon Thereafter: The acquired immune system follows a day or two behind; it shapes the immune response to the type of invader encountered by insuring that specific antibodies are produced.
Mobilize Resources Rapidly: T- and B-cells are the ultimate specialists; only one in perhaps ten thousand can tackle any specific foe. Once activated, these specialists are capable of multiplying rapidly to meet the challenge. Other elements of the immune system (e.g., macrophages) need not multiply to be effective.
Exploit Redundancy: The immune system is highly differentiated and robust against a bewildering array of pathogens. Pathogens, which attack cells and organs throughout the body, come in all shapes and sizes. Some linger in the bloodstream; others d ive for a friendly cellular host at the first opportunity. A robust immune system must take care of all of them, which is one reason that immunoglobulins come in five primary (and many more secondary) flavors, each suited to a particular assignment. The v arious MHC molecules that grip antigens for presentation to the immune cell must sport a variety of holds to match the profusion of potential attackers and their various peptide sequences.
Learn to Act with Minimal Command and Control: The immune system is the epitome of one designed to give actors mission responsibilities to each other, but which thereafter do their job without any further top-down command. It lacks real-time comman d- and-control. Functionality is built into design rather than into an explicit signalling mechanism even though the body cannot anticipate where the next invasion will come from. As with Admiral Nelson's Band of Brothers who intuit their commander's gene ral intent, architecture (and training), rather than fingertip crisis management, is decisive.
Learn to Act with Minimal Information: The immune system operates on a severe economy of inputs. An immune response does not recognize an entire antigen but rather is triggered by individual epitopes[105].
Balance Reaction and Inaction: An immune system which is on all the time is untenable. Even if it has the energy to sustain itself, constant activation risks auto-immune reactions in which too many self proteins are erroneously recognized as antige ns. A targeting system must maintain a balance between tolerating false negative targets (the enemy slips through) or tolerating false positive targets (friends and neutrals are hit); the same holds for winnowing trustworthy from misleading information. T he immune system suggests that a gradation among alert states may be useful for information warfare, so that periods of increasing stress, triggered by suspicious events or other nonself activities, are associated with finely calibrated and multidimension al filters, validation tests, and other protections.
Learn from both Historic and Recent Experience: The innate immune system's reaction to certain proteins and the evolution of MHC cells against specific antigens is akin to learning from history. The presence of memory cells, and retaining APCs to r egenerate them is akin to learning from one's own experiences. The combination is analogous to doctrine that incorporates past lessons but can accept innovation to respond to unforeseen threats.
Balance Concentration and Dispersion: The opposed principles of concentration (the need to couple specific T-cells and B-cells in a cognate reaction in the lymphatic system) and patrol or dispersion (T-cells wandering through the bloodstream) are b oth essential to the immune system.
Remember that Information Warfare Pervades Combat: Much of the human immune system is designed to pass and process information, rather than to apply force. APCs and MHC molecules emulate networks, presenting this or that antigen for inspection by o ne or another T-cell. Helper T-cells function as indications-and- warning systems. Immunoglobulins tag antigens for subsequent destruction. All (but two) of the molecules in the complement chain exist to turn another molecule on. The macrophage has two mi ssions, to eat pathogens and to act as fine-grained molecular sensors. Killer T-cells, which come closest to resembling autonomous warriors, can be activated only by a complex series of checkpoints.
Combining the importance of information warfare with the ability to mobilize suggests that an armed forces composed of specialist corps can be effective -- if methods exist to expand and amplify the core of specialists whose doctrine and capabilities are best suited to defeat a specific foe.
Implications for Information Systems
The immune system may also hold lessons for DOD when it comes time to integrate its various information systems into an overarching network-based framework. Two in particular are knowledge processing and systematic security.
Knowledge Processing: A key component of intelligence-based warfare is the ability to take a heterogenous mass of data and pound some useful conclusions from this. Tomorrow's emerging system of systems may be likened to an enormous inference engine , mobilizing chains of facts to trigger rules which create more facts (e.g., through syllogism, or induction). The immune system, in a sense, is a distributed computer for solving some very complex problems, which (in common with many other biochemical pr ocesses) works through a series of pathways in which one reaction causes the emission of chemicals (and the multiplication of the specifically triggered cell) that spur other reactions. In a system of systems, chaining can be applied to requests for infor mation and the discovery of this information or sources for similar information. If the various agents in a system have receptors for each other, a certain level of random mixing and matching can provide a robust and rapid series of responses to events in troduced into the system. In immune systems, successful reactions often spur replication of the reaction's results; perhaps problems in knowledge representation can benefit by giving disproportionate weight to rules which have worked well in a specific si tuation.
Systemic Security: Can studying human defenses against biology's viruses teach analysts anything about defenses against computer viruses? Most antivirus software depends on an inventory of known viruses to prevent their attack but is helpless again st newly invented ones it does not recognize. Computer researchers have looked at the human immune system to learn how to handle unknown viruses.[106] One approach sets up a dummy program for the virus to infect while a sec ond program analyzes the infected dummy program to locate a characteristic signature of the virus and to communicate this signature throughout the network. Thus the virus can be identified and eradicated. Alternatively, native bit strings can be character ized as self proteins, which are then eliminated from the program's search repertoire.[107]
Another approach to securing complex networks is to develop security agents continually testing computers for the presence of potential antigens -- like the (unproven) theory of immune surveillance, according to which immune cells detect and destroy emerg ing neoplasms.
At the present stage of computer science, large systems tend to be fragile. If the network immune system were, for example, to locate and destroy code that later turned out to be a functioning part of itself, the effect would be more serious than the smal l size of the deleted code might suggest. It is worth noting that those organs of the human body which can least afford bit errors -- brains, eyes, and testicles -- are kept isolated from the human immune system. The brain's immune system does not work on self/nonself distinctions; instead, diseased cells are isolated and disposed of.[108] The analog immune system has built into itself various complex mechanisms to control autoimmune responses, which the digital computer sy stems lack.
Sufficient appreciation of how the immune system works ought to introduce many useful ways to examine military concepts in a new light. The immune systems works as it does because it must respond to particular problems posed in a particular environm ent -- one that is analog, low-bandwidth, and fuzzy. Overall, it seems to be a better metaphor for guiding the use of force in the information fog and the action friction of war; it seems less suited to the digital, high-bandwidth, and mathematical world of cyberspace.
.
.
.
[library]
.
.
.
![]()

![]()
.
.
.
![]()